
Abstract
Environmental monitoring represents an integral part of the microbiological quality control system of a pharmaceutical manufacturing operation. However, guidance documents differ regarding recommendation of a procedure, particularly regarding incubation time, incubation temperature, or nutrient media. Because of these discrepancies, many manufacturers decide for a particular environmental monitoring sample incubation strategy and support this decision with validation data. Such validations are typically laboratory-based in vitro studies, meaning that these are based on comparing incubation conditions and nutrient media through use of cultured microorganisms. An informal survey of the results of these in vitro studies performed at Novartis or European manufacturing sites of different pharmaceutical companies highlighted that no consensus regarding the optimal incubation conditions for microbial recovery existed. To address this question differently, we collected a significant amount of samples directly from air, inanimate surfaces, and personnel in pharmaceutical production and packaging rooms during manufacturing operation (in situ study). Samples were incubated under different conditions suggested in regulatory guidelines, and recovery of total aerobic microorganisms as well as moulds was assessed. We found the highest recovery of total aerobic count from areas with personnel flow using a general microbiological growth medium incubated at 30–35 °C. The highest recovery of moulds was obtained with mycological medium incubated at 20–25 °C. Single-plate strategies (two-temperature incubation or an intermediate incubation temperature of 25–30 °C) also yielded reasonable recovery of total aerobic count and moulds. However, recovery of moulds was found to be highly inefficient at 30–35 °C compared to lower incubation temperatures. This deficiency could not be rectified by subsequent incubation at 20–25 °C. A laboratory-based in vitro study performed in parallel was inconclusive. We consider our results potentially conferrable to other pharmaceutical manufacturing sites in moderate climate zones and believe that these should represent a valuable reference for definition of the incubation strategy of microbiological environmental monitoring samples.
LAY ABSTRACT: Microbiological environmental monitoring confirms that pharmaceutical cleanrooms are in an appropriate hygienic condition for manufacturing of drug products. Guidance documents from different health authorities or expert groups differ regarding recommendation of the applied incubation time, incubation temperature, or nutrient media. Therefore, many pharmaceutical manufacturers perform studies that aim to identify the optimal incubation setup for environmental monitoring samples. An informal survey of the results of such studies, which had been performed at Novartis or European manufacturing sites of different pharmaceutical companies, highlighted no consensus regarding the optimal incubation conditions for microbial recovery. All these studies had been conducted in the laboratory using selections of cultured microbial strains. We tried to solve this disagreement by collecting a significant amount of real environmental monitoring samples directly from the environment in pharmaceutical production and packaging rooms during manufacturing operation. These samples were then incubated under different conditions suggested in the regulatory guidelines. We believe that the results of our study are more meaningful than laboratory-based experiments because we used environmental samples with microorganisms directly isolated from the manufacturing area. Therefore, we believe that our results should represent a valuable reference for definition of the incubation strategy of microbiological environmental monitoring samples.
- Environmental monitoring
- Incubation conditions
- Incubation temperature
- Nutrient media
- Microbiology
- Cleanroom
Introduction
Environmental monitoring (EM) is an integral part of the microbiological quality control system of a pharmaceutical manufacturing operation. The microbiological EM program monitors the overall hygiene status of the manufacturing cleanrooms and determines whether these remain in a predefined state of control. Therefore, the requirement for a scientifically sound EM program is emphasized in several industry guidance documents for sterile manufacturing. However, apart from the expectation that microbiological EM is performed, many of these guidance documents differ regarding recommendation of a procedure. Particularly, the instructions regarding incubation time, incubation temperature, or nutrient media are heterogeneous in both the content as well as in the level of detail. The U.S. Food and Drug Administration (FDA) Guidance for Industry Sterile Drug Products Produced by Aseptic Processing (1), for instance, states that “the microbiological culture media used in EM should be validated as capable of detecting fungi (i.e., yeasts and moulds) as well as bacteria and incubated at appropriate conditions of time and temperature. Total aerobic bacterial count can be obtained by incubating at 30–35 °C for 48 to 72 hours. Total combined yeast and mould count can generally be obtained by incubating at 20–25 °C for 5–7 days”. USP <1116> (2) is less specific considering the incubation time and temperature, but discusses the use of different nutrient media in more detail. Furthermore, the possibility of a two-tiered incubation scheme in which a single tryptic soy agar (TSA) plate is sequentially subjected to a high and low temperature to promote growth of both bacteria and fungi is indicated. It is also interesting to note that both these guidelines disagree regarding their recommendation of the minimum incubation time for total aerobic count: “A general microbiological growth medium such as soybean–casein digest medium (SCDM) is suitable for environmental monitoring in most cases because it supports the growth of a wide range of bacteria, yeast, and moulds. […] If necessary, general mycological media such as Sabouraud's, modified Sabouraud's, or inhibitory mould agar can be used. […] Typically, for general microbiological growth media such as SCDM, incubation temperatures in the ranges of approximately 20–35 °C have been used with an incubation time of not less than 72 hours. Longer incubation times may be considered when contaminants are known to be slow growing. […] In the absence of confirmatory evidence, microbiologists may incubate a single plate at both a low and a higher temperature. Incubating at the lower temperature first may compromise the recovery of Gram-positive cocci that are important because they are often associated with humans”. The Japanese Pharmacopoeia (3) arguably provides the greatest level of detail and differs from both of the incubation times proposed in the guidelines discussed above (1, 2). The general recommendation for detection of aerobes is incubation at 30–35 °C for more than 5 days. For detection of yeasts and moulds, incubation at 20–25 °C for more than 5 days is indicated. The Japanese Pharmacopoeia also explicitly allows an intermediate incubation temperature of 25–30 °C with an incubation period for more than 5 days for recovery of total aerobic count, yeasts and moulds. In addition to guidance regarding incubation temperatures and periods, a selection of different nutrient media is proposed that are considered appropriate for detection of a particular type of contaminant. Finally, the Annex 1 of the EU Guidelines to Good Manufacturing Practice (4) does not provide any guidance regarding incubation conditions. Also, in PDA Technical Report No. 13 (5), which is an industry best practice document, there are no specific recommendations regarding nutrient media and incubation conditions for microbiological EM samples.
As a consequence of this heterogeneous regulatory framework, many manufacturers decide for a particular EM sample treatment strategy and usually support this decision with experimental data in form of a validation. Typically, such validations are performed in the microbiology laboratories as in vitro studies and comprise plating of cultured test microorganisms and their recovery using different incubation conditions and/or nutrient media. The spectrum of microorganisms applied often consists of pharmacopoeial reference strains supplemented with additional species from accredited strain collections as well as in-house isolates. In at least one instance, such a comparative study was also published (6).
Through discussion with representatives from different manufacturing sites of the Novartis group, we noticed that the outcomes of these validation studies were as diverse as the recommendations in the guidelines. Exchange with microbiologists from several European manufacturing sites of different pharmaceutical companies supported a similar conclusion. While some experimentally confirmed the frequently used strategy to recover total aerobic microbial count from TSA incubated at 30–35 °C and total yeasts and moulds count from Sabouraud dextrose agar (SDA) incubated at 20–25 °C, others found different strategies to be at least as efficient or even superior. Applied strategies encompassed the use of only one single TSA plate incubated at an intermediate temperate of 25–30 °C to efficiently recover both bacteria and fungi (as mentioned in Reference 3), or two-tiered incubation schemes as suggested in Reference 2. Some came to the conclusion that incubation of a single TSA plate at 30–35 °C was efficient in recovering both bacteria and fungi, as also was suggested in the literature (6). We saw the large discrepancy in the outcome of these studies, which all had been conducted in vitro using select microbial strains, as indicative of a systematic experimental bias. Therefore, we decided to address this question with a slightly different experimental approach. As opposed to conducting in vitro spiking experiments, we collected real EM samples in pharmaceutical production and packaging rooms of a manufacturing site located in Switzerland and incubated these under different conditions. For the remainder of the article, we will refer to this experimental approach as in situ study. We hypothesized that such an experimental setup would be less affected by strain selection bias, which we suspected to be a potential cause for the variable outcomes of in vitro studies. More detailed considerations regarding the mechanisms by which strain selection bias can influence a study is provided in the discussion section of this paper. As an additional factor, we considered it conceivable that the physiological state and overall fitness of microorganisms at the time of isolation from the environment may be markedly different compared to overnight cultures or other means of preparing an inoculum in the laboratory. Such differences in the physiological state were shown to have an impact on the growth behavior of microorganisms (7). We examined recovery from volumetric air sampling, settling plates, and sampling of inanimate surfaces as well as personnel. In principle, the experimental setup was therefore similar to the one of a recently published study that aimed to compare the recovery efficiency of different mycological media (8). In addition to the in situ study, we addressed the same question in a more conventional way by comparing the same incubation conditions and media in an in vitro study with a selection of 30 different microorganisms.
Materials and Methods
Media
Media used were TSA for recovery of total microbial count and SDA for recovery of total mould count. For volumetric air sampling, settling plates, and in vitro plating studies, 90 mm Petri dishes were used. For sampling from inanimate surfaces and personnel, replicate organism detection and counting (RODAC) plates were used. 90 mm Petri dishes were prepared in-house using dehydrated media from Oxoid (Hampshire, United Kingdom). RODAC plates were purchased from Heipha Dr. Müller GmbH (Eppelheim, Germany) as gamma-sterilized, ready-to-use media supplemented with the neutralizing agents lecithin, Tween 80, histidine and sodium thiosulphate (LTHTh).
In Situ Study
In order to ensure that a sufficient amount and broad spectrum of microorganisms was recovered, in situ samples were derived from pharmaceutical production and packaging rooms of solid, non-sterile drug products corresponding to ISO 8 at rest and their adjacent rooms. Volumetric air samples were taken using MAS-100 air samplers (MBV Ltd., Stäfa, Switzerland), which are based on the impaction method; sampled volume was 1 m3. Settling plates were exposed for 4 h. RODAC plate samples collected from personnel were incubated aerobically or anaerobically; anaerobic conditions were generated by use of the GENbox system (bioMérieux, Marcy l'Etoile, France). Inanimate surfaces sampled were floors and walls; personnel samples were taken from trousers, jackets, sleeves, and the abdominal area of the uniforms of operators working in the production and packaging rooms. All samples were collected over a period of 2 months on a nearly daily basis (excluding weekends). Samples were collected from alternating positions in different rooms and from different operators within these rooms to prevent systematic bias through a microbial population only predominant in a certain room or part of a room. Incubation conditions examined in the main study, which were chosen based on recommendations from guidelines and preliminary data, are indicated in Table I. Incubation conditions of the preliminary study are indicated in Figure 5. Final enumeration of samples was performed after the indicated incubation times; intermediate readings were performed when indicated. Moulds were differentiated from other types of microorganisms based on colony morphology.
Incubation Conditions Examined in the Present Study
In Vitro Study
For the in vitro part of the study, 30 different microorganisms were used (Table II). Microorganisms were either derived from the in-house cryo-culture collection or from an overnight culture and plated onto 90 mm Petri dishes at a concentration of 10–200 CFU (i.e., countable range). Microorganisms of the in-house cryo-culture collection had been grown to late log or stationary phase, transferred into protective medium containing glycerol, and quickly frozen for long-term storage in cryo-tubes at −80 °C. The microorganisms maintained in cryo-tubes were thawed at room temperature immediately prior to use (representing the standard procedure in our laboratories). Overnight cultures had been incubated under conditions defined in preliminary experiments and were diluted to the desired concentration based on determination of the optical density at 600 nm. Incubation conditions and nutrient media used were those described in Table I. Final enumeration of samples was performed after the indicated incubation times; intermediate readings were performed when indicated.
Microorganisms Used for the In Vitro Study
Statistical Analysis
Prism 6.0 (GraphPad Software, Inc.) was used for statistical calculation. For results of in vitro experiments with individual microbial species (Figure 3B, C, and D) normal distribution was assumed. For all other types of experimental data, non-parametric test methods were preferred. These decisions were supported by retrospective use of the Shapiro-Wilk test for normality. With exception of Figure 3A, the significance level of all statistical tests was 5%. In that particular case, the Bonferroni correction was applied in order to account for multiple comparisons, resulting in an adjustment of the significance level to 1.6%.
Results
In Situ Study
Samples of air and inanimate surfaces were taken in pharmaceutical production and packaging rooms during routine operation and incubated under different conditions. At least 70 replicates for each incubation condition were generated by volumetric air sampling, settling plates, and sampling of inanimate surfaces. Volumetric air sampling resulted in the highest mean microbial count numbers followed by settling plates; sampling of inanimate surfaces resulted in the lowest microbial count results. The response to the different incubation conditions, however, was similar irrespective of the sampling method used (not shown). Therefore, results from the different sampling methods were pooled in order to increase sample size to at least 210 replicates per incubation condition.
Recovery of total aerobic microorganisms seemed to be numerically superior at temperatures 25–30 °C and 30–35 °C as compared to two-tiered incubation conditions starting with 20–25 °C (Figure 1A). This difference was not statistically significant with the used sample size (P = 0.06); however, a trend towards systematically lower numerical recovery was evident. Obtained mean microbial count after incubation at 30–35 °C for 3 days was nearly identical to incubation at 25–30 °C for 7 days. The absolute number of recovered moulds was low. However, differences between the incubation conditions of interest with respect to the recovered number of mould colonies became evident through the large sample size. The number of detected mould colonies was the highest on SDA incubated for 7 days at 20–25 °C (Figure 1B). On average, two-tiered incubation on TSA starting with 20–25 °C recovered approximately 75% of mould count when compared to incubation at 20–25 °C on SDA, which was not statistically significant with the used sample size (P = 0.19). Incubation on TSA at 25–30 °C for 7 days recovered approximately 45% of mould count compared to incubation at 20–25 °C on SDA, which was significantly less. Incubation at 30–35 °C for 3 days yielded the lowest number of mould colonies and was significantly worse than any other incubation condition tested; when compared to incubation at 20–25 °C on SDA, recovery was approximately 10%.

Microbial recovery from EM samples incubated under different conditions. Recovery of total aerobic microorganisms (A) and moulds (B) is shown. Mean value and standard error of the mean (SEM) are indicated from at least 210 replicates per incubation condition. Two-tiered incubation means incubation at 20–25 °C for 5–6 days followed by 2–3 days at 30–35 °C. If not otherwise indicated, used nutrient medium was TSA. Significant differences in CFU count were calculated with Kruskal-Wallis test and Dunn's test corrected for multiple comparisons as post-test (ns, not significant; *, P < 0.05; ****, P < 0.0001).
Samples from personnel were taken from operators working in pharmaceutical production and packaging rooms. Samples were either incubated aerobically or anaerobically; at least 50 replicates were generated for each incubation condition. For recovery of total aerobic microorganisms, superior count results were obtained with incubation at 30–35 °C for 7 days as compared to two-tiered incubation conditions starting at 20–25 °C (Figure 2A). This was similar to the results observed for air and inanimate surfaces (Figure 1A), although the magnitude of the effect was clearly more pronounced for the personnel samples. A similar picture emerged for anaerobic incubation conditions (Figure 2B).
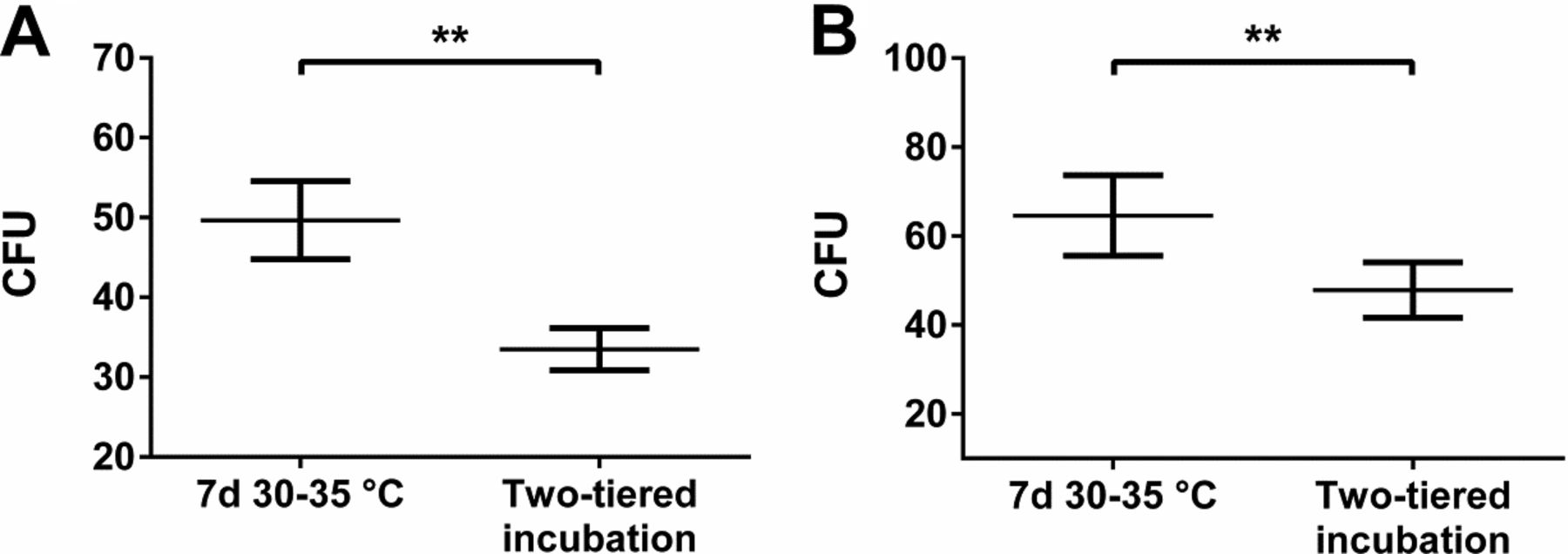
Microbial recovery from personnel monitoring samples incubated under different conditions. Recovery of total aerobic microorganisms (A) and total anaerobic microorganisms (B) is shown. Mean value and standard error of the mean (SEM) are indicated from at least 50 replicates per incubation condition. Two-tiered incubation means incubation at 20–25 °C for 5–6 days followed by 2–3 days at 30–35 °C. Used nutrient medium was TSA. Significant differences in CFU count calculated with Mann-Whitney test are indicated (ns, not significant; **, P < 0.01).
In Vitro Study
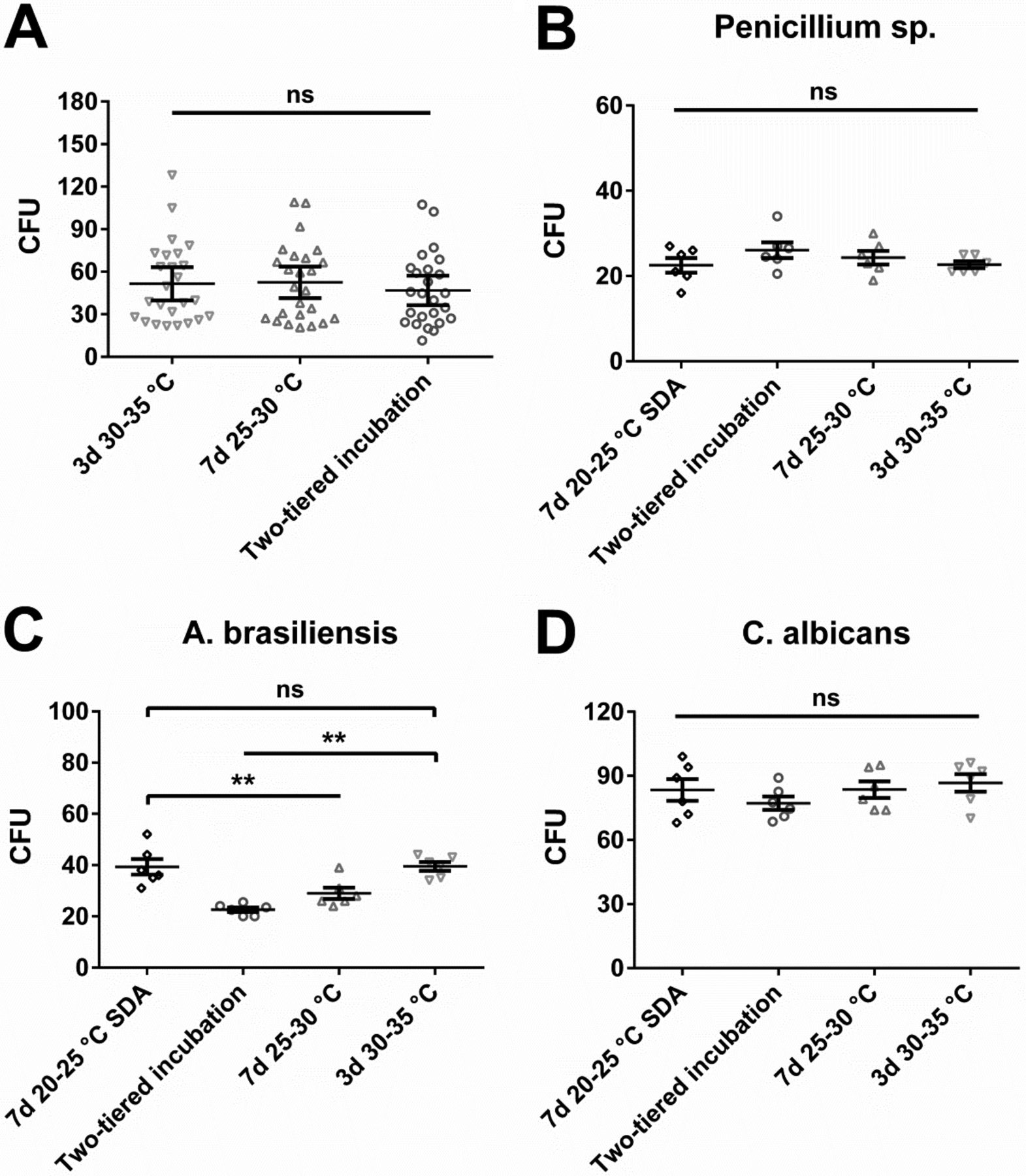
We decided to compare results obtained during in situ sampling to a laboratory-based in vitro study. Such in vitro studies represent a frequently used approach to validate incubation conditions for EM samples. We defined a microbial spectrum consisting of 30 different microbial strains from our cryo-culture collection (Table II). The chosen microorganisms consisted of pharmacopoeial reference strains as well as in-house isolates, which had been collected from the environment or been recovered in product release tests in the past. Microorganisms were grouped based on their expected relevance for EM or personnel monitoring, as well as their ability to grow under aerobic or anaerobic conditions. Using the conditions examined for incubation of air and inanimate surface samples, no differences were detectable for bacteria (Figure 3A). In contrast to the in situ samples (Figure 1A), no tendency towards superiority of a particular incubation condition was observable as visualized by a complete overlap of the error bars of the mean CFU values. Moreover, application of Wilcoxon's matched-pairs rank sum test, which compared the mean values obtained for each individual microbial strain in a pairwise manner, did not result in a significant difference. Interesting results were obtained for the used fungal strains (Figure 3B, C, D). In sharp contrast to the in situ data (Figure 1B), incubation at 30–35 °C for 3 days appeared as a viable option for mould detection. Aspergillus brasiliensis, the pharmacopoeial reference mould strain, was recovered significantly better at 30–35 °C as compared to lower incubation temperatures when TSA was used as nutrient medium; incubation at 20–25 °C on SDA for 7 days led to similarly efficient recovery. The in-house mould isolate Penicillium sp. as well as the yeast Candida albicans did not exhibit a clear preference.
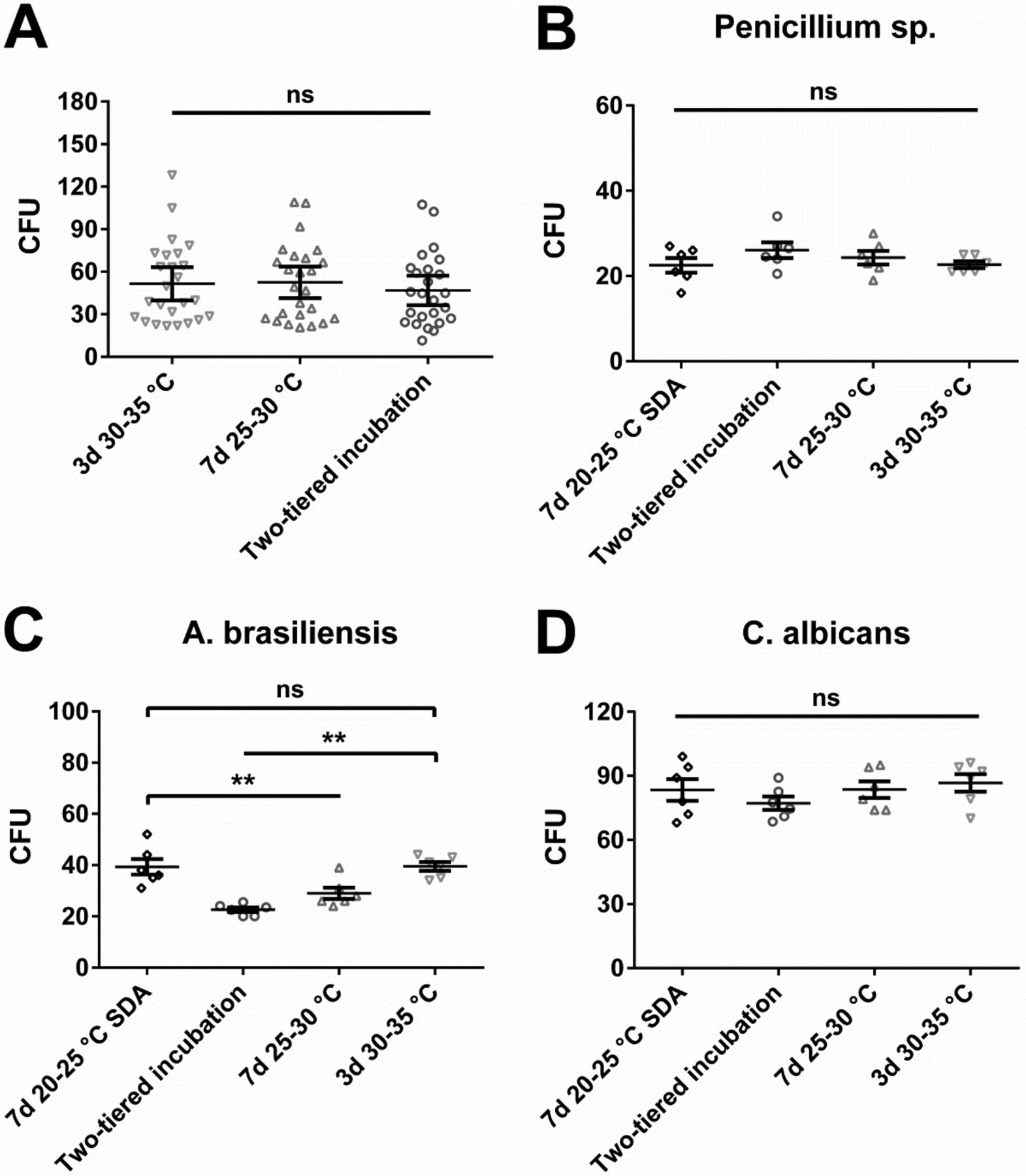
In vitro study with cryo-cultured microorganisms incubated under different conditions evaluated for EM (air and inanimate surfaces). Two-tiered incubation means incubation at 20–25 °C for 5–6 days followed by 2–3 days at 30–35 °C. If not otherwise indicated, used nutrient medium was TSA. (A) Recovery of 25 different bacteria. Each symbol represents the mean CFU value of a particular microorganism obtained from at least six replicates; mean of these values and standard error of this mean (SEM) are indicated. No significant difference with Wilcoxon's matched-pairs signed rank test (values for each microorganism at each incubation condition were paired for the calculation and the difference between these paired values assessed). (B, C, D) Recovery of three different fungi; each symbol represents an individual replicate. Furthermore, mean value and SEM of these replicates is indicated. Statistically significant differences were calculated with one-way analysis of variance (ANOVA) and the Tukey-Kramer method as post-test (ns, not significant; **, P < 0.01).
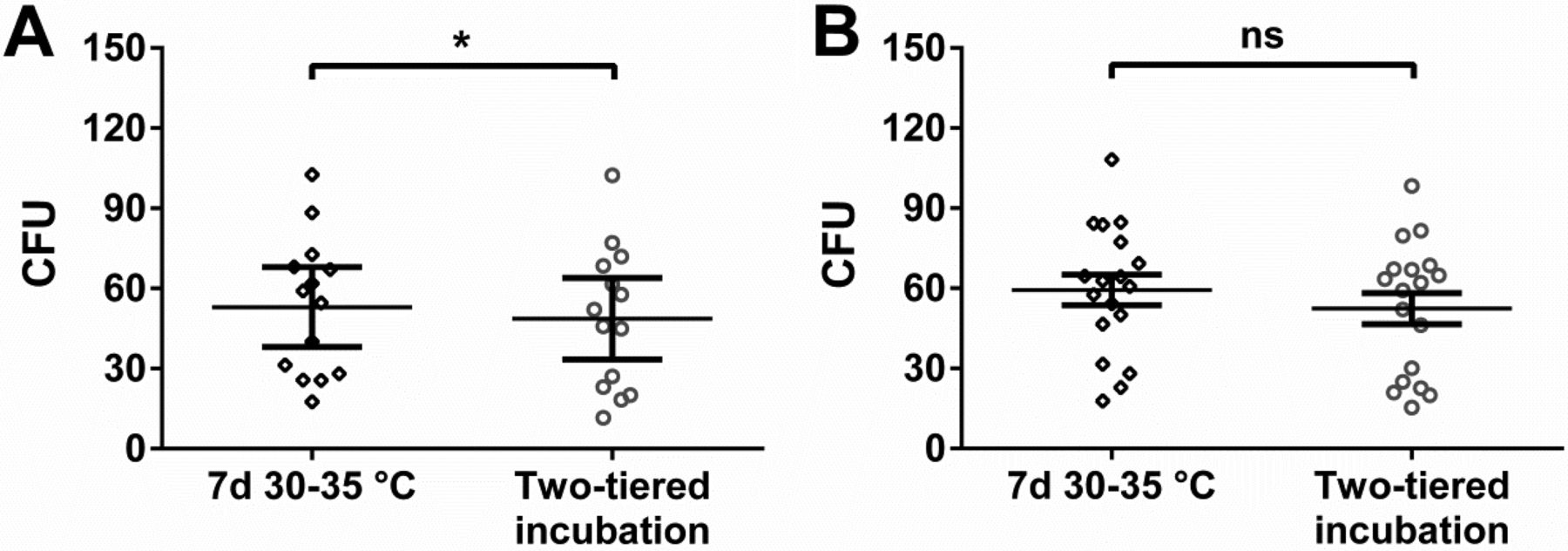
Using the incubation conditions examined for personnel monitoring samples, no evident differences in mean microbial recovery were detectable (Figure 4A, B). Application of Wilcoxon's matched-pairs rank sum test resulted in a significant difference in the favor of incubation at 30–35 °C with the aerobic, but not the anaerobic, incubation conditions. However, differences were less pronounced if compared to the results obtained in situ (Figure 2).
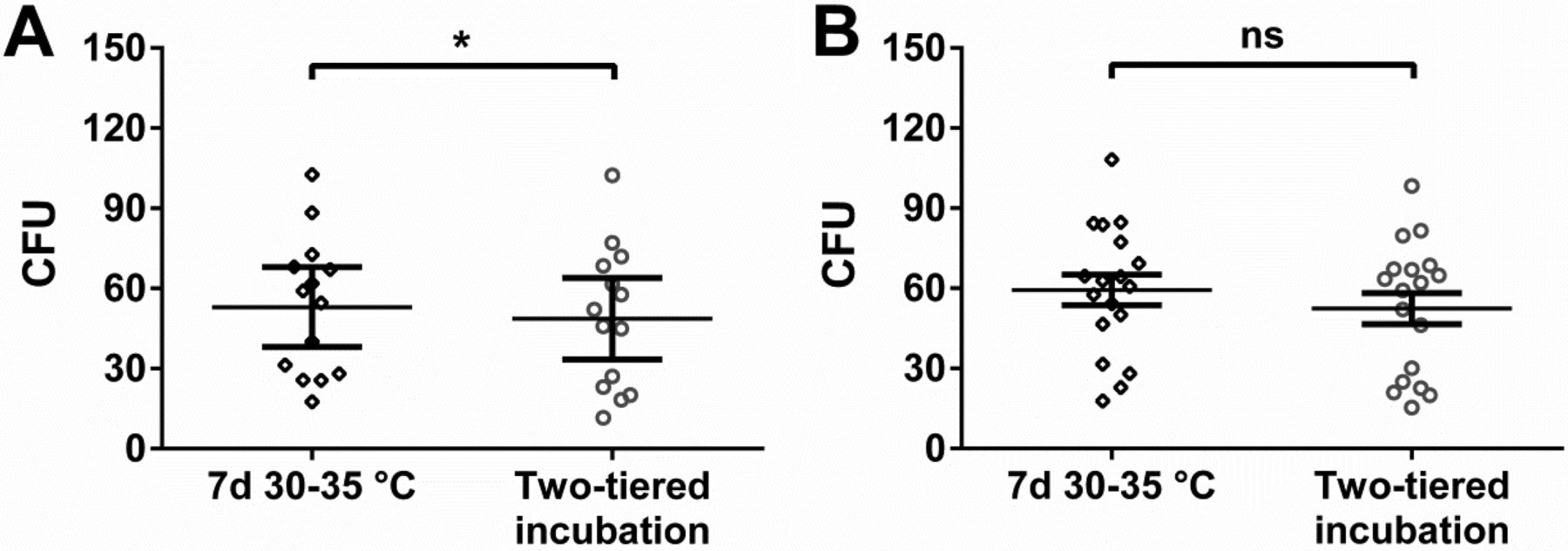
In vitro study with cryo-cultured microorganisms incubated under different conditions evaluated for personnel monitoring. Two-tiered incubation means incubation at 20–25 °C for 5–6 days followed by 2–3 days at 30–35 °C. Used nutrient medium was TSA. (A) Recovery of 14 different bacteria incubated under aerobic conditions and (B) 18 different bacteria incubated under anaerobic conditions. Each symbol represents the mean CFU value of a particular microorganism obtained from at least six replicates; mean of these values and standard error of this mean (SEM) are indicated. Statistically significant differences with Wilcoxon's matched-pairs signed rank test are indicated (ns, not significant; *, P < 0.05). Values for each microorganism at each incubation condition were paired for the calculation and the difference between these paired values assessed.
Discussion
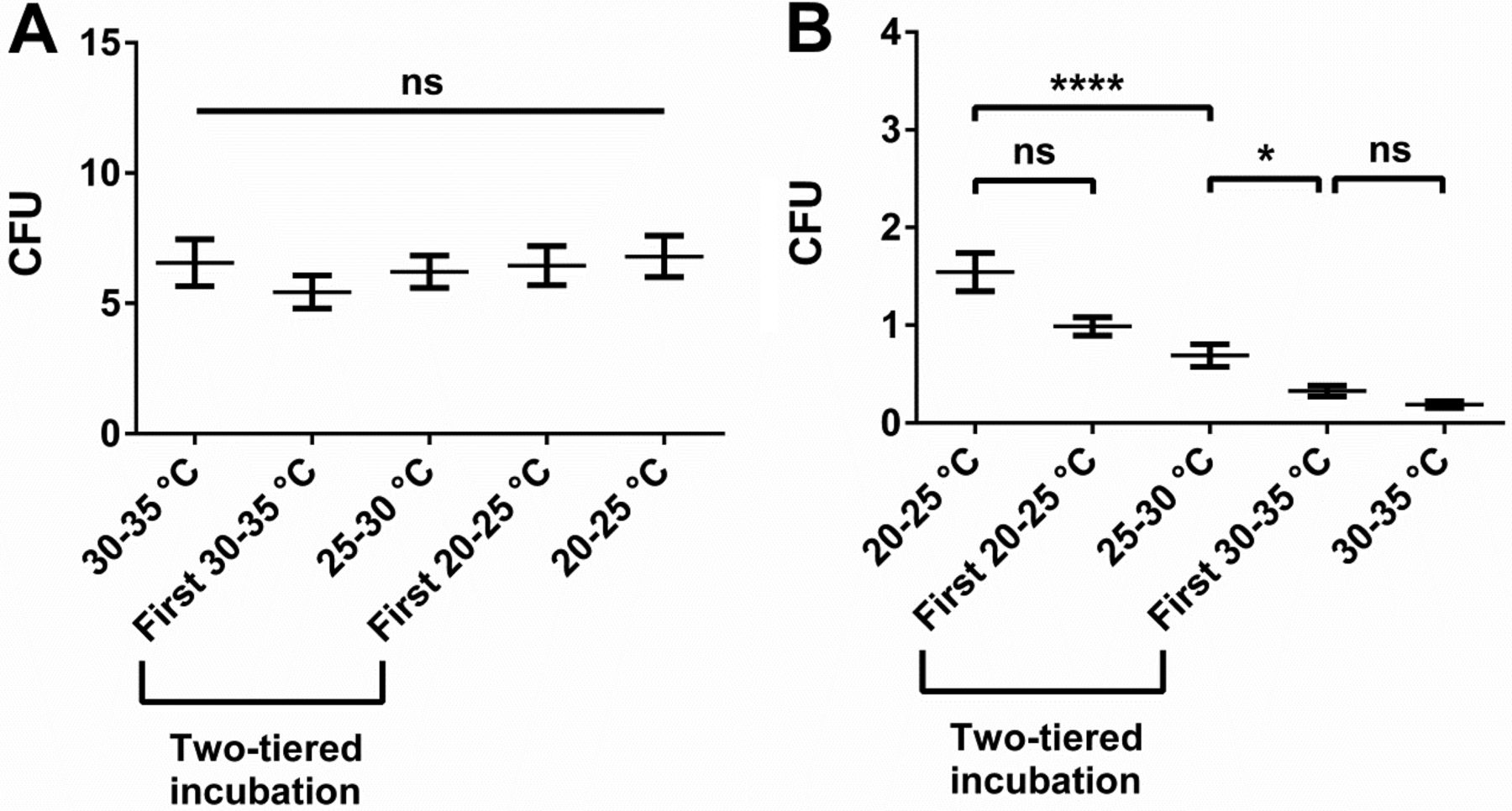
In our in situ approach we observed distinct differences in recovery of total aerobic microbial count or mould count between the incubation conditions of interest. Recovery of total aerobic microbial count was most efficient at elevated temperatures, particularly if samples were taken from gowns of personnel (Figure 2A). We considered this observation reasonable—most microorganisms on gowns are rather derived from personnel than the environment, and microorganisms colonizing humans should be adapted to preferentially grow at temperatures close to the skin temperature of approximately 35 °C (9). The observation that total aerobic count from air and inanimate surfaces also showed a trend towards superior recovery at elevated temperatures may seem less straightforward (Figure 1A). Microorganisms naturally occurring in the environment are generally not expected to prefer elevated temperatures for replication because such temperatures are not frequently encountered in a central European climate zone. For instance, microbial recovery from water, which can be a source of microorganisms in a cleanroom (10), was shown to be more efficient at lower temperatures (11). Inhibited growth of moulds at elevated incubation temperatures, and thus less overgrowth of the bacterial colonies, may offer a possible explanation. However, the intermediate readings throughout the incubation period probably mitigated such an effect. Furthermore, the low numbers of recovered moulds suggest that overgrowth had no significant contribution to this observation. Alternatively, the observed preference is probably attributed to the fact that a good proportion of microorganisms in a pharmaceutical cleanroom is actually shed by personnel (12, 13). Further support for this explanation can be derived from a preliminary study we conducted at our site (Figure 5). Whereas the samples of the main study originated from production and packaging rooms that were in operation, samples of the preliminary study were collected in a secluded area without personnel flow. In the preliminary study, no indication for superior recovery of total aerobic microbial count at 30–35 °C as compared to lower incubation temperatures was observable (Figure 5A), whereas such a trend was evident in the main study (Figure 1A). Thus, the presence of personnel and the resulting shedding of human-associated microorganisms in the environment may have been the underlying cause for the superior recovery of total aerobic microbial count from air and inanimate surfaces at elevated temperatures.
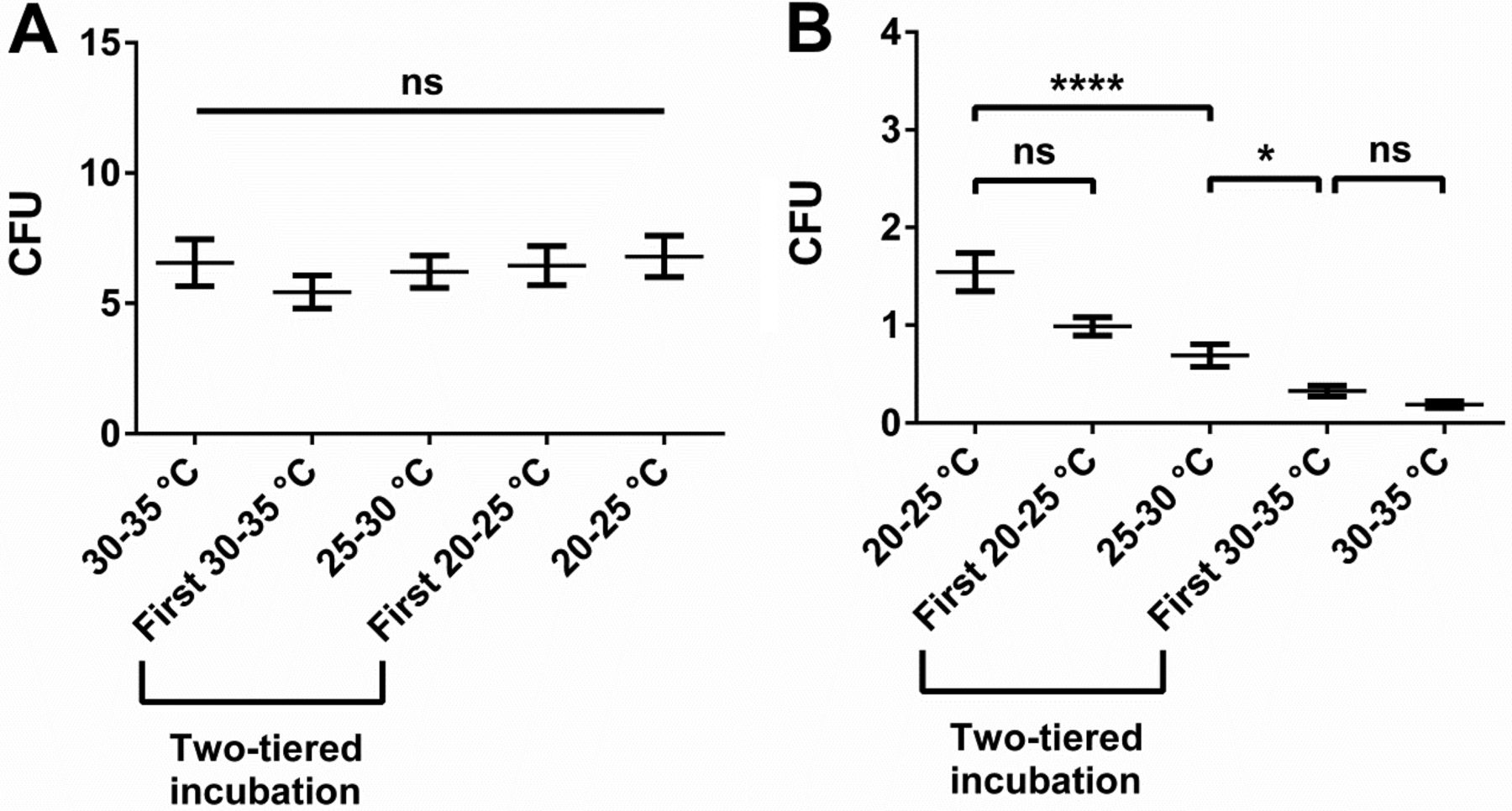
Microbial recovery results from a preliminary in situ study. Data was collected in a secluded location without personnel flow. Total incubation time was 5 days and used nutrient medium was TSA. Recovery of total aerobic microorganisms excluding moulds (A) and moulds (B) is shown. Moulds were subtracted from total aerobic microorganisms count because of their untypically high relative abundance in this experiment and thus strong influence on the total aerobic microorganism count result. Mean value and standard error of the mean (SEM) are indicated from at least 170 replicates per incubation condition. Two-tiered incubation means incubation for 3 days at 20–25 °C and for 2 days at 30–35 °C; the initial incubation condition is specified. Significant differences in CFU count were calculated with Kruskal-Wallis test and Dunn's test corrected for multiple comparisons as post-test (ns, not significant; *, P < 0.05; ****, P < 0.0001).
In contrast to total aerobic count, recovery of moulds continuously and drastically decreased with increasing incubation temperatures (Figure 1B and 5B). Also this observation seemed plausible because indoor moulds in a temperature-controlled environment have only weak selective pressure to be able to efficiently grow at temperatures above 25 °C. Initial incubation at 30–35 °C seemed to severely impair mould growth despite subsequent incubation at 20–25 °C (Figure 5B), arguably challenging that incubation with the high temperature first represents a generally superior approach for two-tiered incubation schemes as some may infer from USP <1116> (2). Whether mould recovery could be increased through prolonged incubation at 20–25 °C after being subjected to 30–35 °C should be a topic for further research. The use of the mycological medium SDA conveyed a slight increase in mould recovery compared to TSA. Of interest, in a recent retrospective study in hospital pharmacies and compounding pharmacies, Weissfeld et al. came to the conclusion that the use of mycological media (in this case yeast malt extract agar) is important (14). However, in the study by Weissfeld et al. the mycological medium was incubated at 28 ± 2 °C and the general microbiological medium (TSA supplemented with polysorbate and lecithin) was incubated at 35 ± 2°C as recommended in one of their relevant guidelines, USP <797> (15). Thus, the study does not allow dissecting the contribution of incubation temperature or nutrient medium towards the observed difference in fungal recovery. Our study highlights that the differences observed in the study by Weissfeld et al. may also be explained by a temperature effect rather than a nutrient medium effect (Figures 1B and 5B). Furthermore, our data also indicate that the incubation temperature of 28 ± 2 °C proposed for mycological media in USP <797> (15) may arguably be too high for optimal recovery of environmental moulds.
The differences observed in the in situ study were only poorly reflected in the in vitro study despite application of a strain-specifically paired statistical evaluation approach. We hypothesize that the underlying cause of this observation was the strain selection. Although some of the used microorganisms clearly preferred a particular incubation condition, these preferences leveled out with the microbial selection used for this study. With another selection of microorganisms, however, a different answer may have been obtained. Strain selection bias may probably be the main cause for the very different outcomes of EM validation studies performed at different laboratories and companies. In general, we see four main contributors to strain selection bias.
First, the most prominent effect of incubation conditions on microbial recovery from air and inanimate surfaces was observed for moulds. However, in a well-controlled environment, moulds only represent a small fraction of microorganisms recovered from EM samples (particularly in higher grade cleanrooms in sterile manufacturing) (13). This may elicit a tendency to represent moulds less well in study designs. Our own in vitro study, which had been defined based on microorganisms available in our cryo collection, was strongly affected by this deficiency.
Second, the pharmacopeia-referenced A. brasiliensis mould strain (ATCC 16404) has an obvious preference for elevated incubation temperatures (Figure 3C). As demonstrated in the present study, this preference seems to deviate from the temperature range optimal for recovery of moulds from the environment. Thus, this strain may pose a questionable reference with regards to EM. The other pharmacopoeia-referenced strains which were included in our study did not exhibit clear temperature preferences (not shown). Obviously, most of the pharmacopoeia-referenced strains can efficiently be recovered within a broad temperature range, and as no clear preferences exist, a variety of different incubation conditions could be defined on basis of these results. However, whether this broad temperature tolerance is also representative for the majority of environmental microorganisms cannot be deduced. Thus, it should be carefully considered which conclusions may validly be drawn for the incubation conditions of the routine monitoring program on basis of such laboratory results.
Third, the origin of externally purchased isolates used in a study should be carefully considered. Microbial strains from accredited microorganism collections often represent patient isolates. The prerequisite for such pathogenicity is the ability or even preference to grow at elevated temperatures—but whether such strains are representative for a pharmaceutical manufacturing environment is questionable. Thus, the use of such strains may lead to a bias towards preference of high incubation temperatures, although this may not reflect the preference of most environmental strains. An earlier study claiming that an incubation temperature of 30–35 °C is ideal for fungal detection (6) featured a microbial spectrum in which four of nine strains were patient isolates. This strain selection bias at least partially contributed to a study outcome not supportable by data generated from actual environmental samples.
Finally, in the case of in-house isolates, the incubation condition used for isolation is also a potential source of significant bias. For instance, if the EM program used to date consisted of incubation at an elevated temperature, the sole reliance on isolates obtained at that temperature is problematic. This is due to the fact that microorganisms not able to grow at these elevated temperatures would not have been isolated. Therefore, the isolate selection would be highly biased towards confirmation of the previously used incubation conditions. Our in-house isolate Penicillium sp., for instance, had been isolated at more than 25 °C; thus, it is not surprising that this particular strain could tolerate elevated incubation temperatures (Figure 3B). Accordingly, even if we had included a larger number of mould isolates from our environment in the in vitro study, the significance of these results would remain questionable. Because the incubation temperature routinely applied to that date had been 25–30 °C, there is a very high probability that an in vitro study with moulds isolated from routine EM would confirm these conditions—although this may not necessarily be representative for the actual environment. Similarly, if no mycological media had been used in the past, there is a high probability that a validation applying previously isolated fungi would not point towards a superior recovery with mycological media as compared to the general microbiological growth medium used for isolation—based on the fact that fungi that indeed strongly prefer mycological media had a lower chance to be included in the isolate collection. Therefore, in order to have a strain selection which is less affected by bias, isolation would have to occur using all of the different media and incubation temperatures of interest, which essentially resembles an in situ approach.
As outlined above, introduction of bias in in vitro experiments aiming towards comparison of different EM incubation conditions is difficult to avoid. Therefore, we consider the results of our in situ approach as more meaningful. We also believe that our study may be considered generally representative for manufacturing sites in similar climate zones. Although the exact strain composition at different sites may differ, temperature preferences of the indigenous microorganisms should remain comparable. For instance, microorganisms of human origin may be expected to be better recoverable at temperatures close to the skin temperature (as also supported by the data in Figure 2A and B). Accordingly, if it is assumed that the predominant part of the cleanroom microflora in a particular area is human-derived, one could also reasonably expect superior recovery of environmental microorganisms at elevated incubation temperatures. Our results obtained for an area with personnel flow (Figure 1A), in which such a trend was evident, and an area without personnel flow (Figure 5A), where no correlation between incubation temperature and recovery of total aerobic microorganisms was observable, support this notion. Such factors should, in principle, be independent of the manufacturing site. However, whether a generalized shift towards a preference of higher incubation temperatures (particularly for moulds) would be observed in cleanrooms located in a tropical climate zone may be a question to be further elucidated. The observation that fungal recovery from tropical forest soil was superior at an incubation temperature of 30 °C as compared to 25 °C indicates that this may indeed be the case (16).
In our study design we to a certain extent omitted yeasts, which were only reflected in total aerobic count. The reason for this apparent shortcoming was the expected difficulty to unambiguously differentiate some yeast from bacteria based on colony morphology. The large sample size of our study forbade systematic screening of all recovered colonies with a microscope or use of other identification techniques. In principle, recovery of yeasts from the environment could follow a similar pattern regarding impact of different temperatures and nutrient media as was observed for moulds. This assumption may be supported by the view expressed in several guidelines that recommend that the same incubation conditions and nutrient media should be used for detection of yeasts as well as moulds (1, 3). However, the authors are not aware of studies experimentally addressing that issue for pharmaceutical cleanrooms. An obvious way to elucidate this question would be an assessment of the impact of the different temperatures and nutrient media in an in situ study using SDA and TSA supplemented with antibiotics.
Based on our results, the highest overall recovery from aerobic environments (air, inanimate surfaces) would have been achieved when total aerobic count was obtained from TSA incubated at 30–35 °C and mould count from SDA incubated at 20–25 °C. But also other approaches yielded acceptable results. While the two-tiered incubation strategy had a slight deficiency in recovering total aerobic count, mould detection was very effective. Incubation at an intermediate temperature of 25–30 °C, on the other hand, led to very efficient recovery of total aerobic count, but had certain deficiencies in recovering moulds. However, when defining an incubation strategy for EM samples, it should be emphasized that the use of two plates with different nutrient media is not optimal in every case. For monitoring of an aseptic manufacturing operation, for instance, the double-sampling dictated by use of two nutrient media would lead to a higher number or an extended duration of interventions or manipulations close to open product as compared to a single-plate strategy. This potential concern may be less relevant when production occurs in an isolator or involves use of a restricted access barrier system. Moreover, human-derived bacteria typically represent by far the most common contaminant in well-controlled ISO 4.8/5 environments, whereas fungi can be considered rare (1%) in comparison (13). These considerations further strengthen the argument that monitoring of such areas should not regularly be performed with two media, as the higher product contamination risk through the operator may outweigh the slightly improved capability to detect fungi. Rather, a monitoring strategy that relies on a single nutrient medium incubated under conditions allowing for reasonable detection of bacteria, yeasts, and moulds (i.e., intermediate temperature of 25–30 °C or two-tiered incubation) complemented with periodic use of mycological media may represent a more appropriate approach. It is also worth mentioning that the above rationale supports the recommendation provided in USP <1116> (2) to initially apply the high temperature for two-tiered incubation schemes in order to efficiently recover human-associated bacteria. However, the resulting recovery of moulds seems to be so low (Figure 5B) that the value of the subsequent change to a reduced incubation temperature should be addressed in systematic studies. Eventually, an extension of incubation time at the reduced temperature may ameliorate mould recovery. Also, further work focused on yeasts may additionally support or oppose the necessity of the proposed temperature shift.
Similar to ISO 4.8/5 environments, we believe that a single-plate strategy may also allow for appropriate surveillance of the microbiological state of control of areas with higher microbial burden. In particular, the definition of appropriate alert levels and data trending strategies enable assessment of the microbiological control status irrespective of whether the overall microbial recovery would be higher through the use of two nutrient media. However, in areas where product considered distinctly critical regarding fungal contamination (e.g., some inhalation products) is openly handled, the use of mycological media at defined frequencies should indeed be considered as part of the EM program in a risk-based manner.
In conclusion, we assessed microbial recovery from EM samples in an in situ approach and compared the results to an in vitro study. Our results indicated highest recovery of total aerobic count in areas with personnel flow using a general microbiological growth medium (TSA) incubated at 30–35 °C and highest recovery of moulds with a mycological medium (SDA) incubated at 20–25 °C. However, also single-plate strategies applying a general microbiological growth medium either in a two-temperature incubation scheme starting at 20–25 °C or using an intermediate incubation temperature of 25–30 °C yielded reasonable results. Recovery of moulds was inversely correlated with the applied incubation temperature. Interestingly, subsequent shift from a high incubation temperature to a low incubation temperature did not seem to ameliorate mould recovery with the incubation time applied. We showed that the in situ approach was more powerful in detecting differences between the incubation conditions of interest. Furthermore, we offered explanations why in vitro studies conducted by different laboratories and companies came to conflicting results and sometimes also counterintuitive conclusions. For instance, the assertion that for recovery of environmental fungi in moderate climate zones incubation above 30 °C should be equivalent to incubation at ambient temperature (6) is conclusively rejected by our study. Indeed, care must be taken when designing laboratory-based in vitro studies in order to mitigate bias, and the extent to which such studies allow investigators to draw conclusions for EM samples should be well reflected. In general, we consider our in situ approach as being more meaningful compared to in vitro studies when examining different incubation conditions for EM samples, and we believe that the results may be potentially conferrable to pharmaceutical manufacturing sites in similar climate zones. Therefore, our study should represent a valuable reference for definition of the incubation strategy of microbiological EM samples.
Conflict of Interest Declaration
The authors declare that they have no competing interests.
Acknowledgments
We thank Eileen Albrecht for collection of in situ and in vitro data and Melanie Nast for supporting her. Catharina Nord is acknowledged for collecting the data of the preliminary study.
- © PDA, Inc. 2014



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Establishment of a Single Temperature Incubation Approach for Environmental Monitoring Samples with Focus on Mold Recoveries
- Multisite Qualification of an Automated Incubator and Colony Counter for Environmental and Bioburden Applications in Pharmaceutical Microbiology
- Environmental and Personnel Monitoring Programs--A Risk-Based Case Study of Cutibacterium acnes
- Mold Control and Detection in Biological Drug Substance Manufacturing Facilities: An Industry Perspective
- Evaluation of the Recovery Rate of Different Swabs for Microbial Environmental Monitoring
- Multicenter Study on Incubation Conditions for Environmental Monitoring and Aseptic Process Simulation